Esempio: consideriamo tre geni w,y, m. Se i geni sono disposti in una struttura lineare allora uno di essi deve trovarsi in una posizione centrale paragonata a quella degli altri due. La distanza genetica più grande quella che separa i geni più esterni e questo valore dovrebbe essere uguale alla somma delle distanze che separano il gene centrale da ciascuno dei geni laterali.
La mappatura per ricombinazione sembrava dare ragione all'ipotesi che i geni sono disposti su una struttura "lineare".
Sturtevant stabilì un ordine non contradditorio di tutti i geni da lui studiati sul cromosoma X, controllando i dati per ogni combinazione di tre geni, ci si rende conto che qusto ordine aveva una logica. Inoltre i dati sulla ricombinazione erano a favore della teoria che si potesse costruire una mappa in cui i geni erano disposti linearmente lungo il cromosoma.

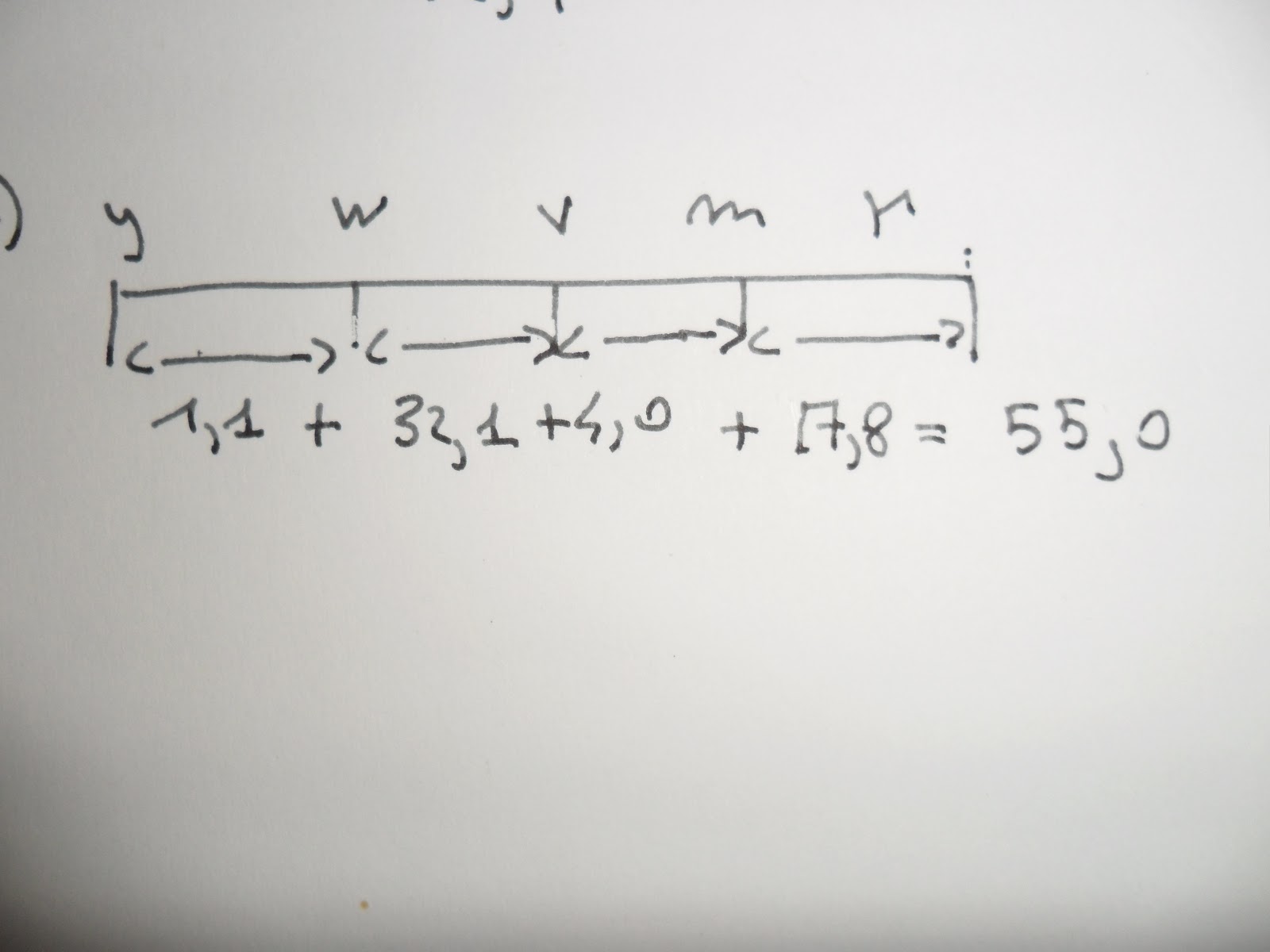
Per esempio, mappando y, m, w, la distanza tra i due geni esterni y e m è 34,3 u.m, la distanza tra i geni entrale w e l'esterno m è circa dello stesso ordine di grandezza 32,8 u.m.; come possiamo notare i valori sono molto simili, per evitare di trovarci difronte ad un errore di campionamento, bisognerebbe incrociare un grandissimo numero di moscerini e sottoporre i dati ad un test statisctico, come il test del chi quadrato.
Inoltre n altro problema che si incontra con il metodo di mappatura di Sturtevant è che le distanze effettive nella mappa non sono sempre addittive.
Esempio: se il locus del gene y si trova all'estrema sinistra della nostra appa n una posizione che possiamo definire come punto zero; il gene w sarebbesituato allora vicino alla posizione 1, ed m nelle vicinanze della posizione 34,2 u.m.
Per il gene r basandoci su quella che è la distanza da y (osserviamo la figura a) dovrebbe trovarsi in posizione 42,9 u.m. il problema sorge quando andiamo a sommare la distanza intermedia, cioè addizionando le distanze y<-->w più la distanza w<--->v più la distanza v<--->m più la distanza il locus r dovrebbe trovarsi ad una distanza di circa 55 u.m. Quale valore tra i due è più vero?
l'incrocio a tre punti può darci qualche risposta a riguardo
Incrocio a tre punti.
Di cosa si tratta questo tipo di incrocio? Nell'incrocio a tre punti analizziamo simultaneamente tre geni, i nostri marcatori, l'utilizzo dell'incrocio a tre punti ci permette di dAnalizzando simultaneamente tre marcatori, possiamo ottenere più facilmente una serie di informazioni che ci permettono di identificare l'esatta posizione dei geni.
Esempio: femmina omozigote per i caratteri ali vestigiali (a); corpo nero (b) occhi gialli (c) viene incrociata con maschio di tipo selvatico con geni a+ b+ c+.
La progenie F1 è eterozigote e mostra, sia per i maschi che per le femmine, un fenotipo normale per i tre caratteri indicando che le mutazioni sono autosomiche e recessive, la progenie F1 è rappresentata da individui tutti identici.

Noi non conosciamo l'ordine dei geni, potrebbe essere a b c o qualsiasi altra combinazione derivante ed indicare lo stesso genotipo, cioè non conoscendo il modo in cui tali geni sono disposti sul cromosoma, noi potremmo anche scrivere b a c oppure c a b ed indicare lo stesso genotipo e fenotipo.
Lo scopo di mappare però è quello di riuscire a comprendere in che ordine sono disposti i geni di nostro interesse su un cromosoma.

Ora prendiamo in considerazione due geni alla volta, numero questo fondamentale per eseguire il calcolo della frequenza di ricombinazione.
Prendiamo la coppia di geni a e b. Le combinazioni parentali sono a+ e b+; i ricombinanti sono a e b+; a+ e b.
Per determinare se una classe particolare della progenie è ricombinante o parentale per queste due classi di geni non importa sapere se i nostri moscerini sono c+ o c.
La distanza a<-->b calcolata come la percentuale di ricombinanti sul numero totale della progenie sarà di 350+338+231+218/5109 x 100= 22,5 u.m

350+338+15+7/5109 x 100= 13,89 u.m
Stesso discorso anche per b e c:
231+218+15+7/5109 x 100= 9,3 u.m
Ciò dimostra che a e b sono la coppia di geni più distanti e di conseguenza possiamo assumere che sul cromosoma sono i geni presenti in posizione più esterna, con il gene c che è localizzato tra questi due geni, quindi in una posizione che potremmo definire "centrale".
Anche in questo caso però possiamo riscontrare lo stesso problema analizzato nell'incrocio a due punti; la distanza tra i geni più esterni non corrisponde alla somma delle distanze intermedie, infatti 13,89 + 9,3 = 23,20 u.m.
Il motivo? Dobbiamo considerare i doppi crossing over!
Prendiamo sempre in considerazione questi tre geni ,quelli che ho provato a disegnare, sono cromosomi autosomi omologhi eterozigoti per i geni di cui abbiamo accennato sopra.
Osservando l'immagine possiamo notare quali sono le regioni coinvolte nel crossing over che hanno dato origine alla progenie osservata precedentemente.
La progenie che avevamo ottenuto in un testcross tra femmine della F1 triple eterozigoti e maschi selvatici omozigoti per alleli recessivi di tutti e tre i caratteri è costituita da otto gruppi.
Ogni gruppo è costituito da un numero differente di moscerini; i due gruppi più numerosi hanno la stessa configurazione genica dei loro nonni, la generazione P (a b c e a+ b+ c+) questi geni costituiscono la classe parentale.
I gruppi successivi (a+ b c e a b+ c+) sono le classi ricombinanti che rappresentano i prodotti reciproci di un crossing over nella regione 1 tra a e c.


In questi due gruppi si sono verificati degli eventi di doppi crossing over differenti avvenuti contemporaneamente, uno nella regione 1 del cromosoma l'altro nella regione 2.

Bisogna tenere conto di questo doppio scambio, infatti la distanza che abbiamo precedentemente calcolato non ne teneva conto , quindi bisogna correggere l'errore nel calcolo della frequenza di ricombinazione aggiungendo a tale calcolo i doppi crossing over, dal momento che ogni individuo che mostra un genotipo tipico del doppio crossing over è il risultato di due scambi tra a e b.
350+338+231+218+15+15+7+7/5109 x100= 23,16
questo valore ora risulterà più corretto, se andiamo a fare i conti e sommiamo anche le distanze intermedie tra i due geni ci renderemo conto che la distanza tra a e b è identica alla somma della distanza tra a e c e tra c e b.
Quando Sturtevant inizialmente mappò i geni mediante incroci a due punti, la posizione del locus per il gene delle ali rudimentali risultava ambigua. Un incrocio a due punti dava una frequenza di ricombinazione di 42,9 mentre la somma delle distanze intermedie dava un valore più elevato, 55,0.
Questo era dovuto al fatto che la mappa a due punti ignora il processo del doppio crossing over avvenuto nel vasto intervallo dei geni y e r. Invece sommando i valori di distanze più piccole ciò può essere notato più facilmente anche perchè un evento di doppio crossing over difficilmente avviene in intervalli molto piccoli.
Bisogna sottolineare però che anche una mappa a tre punti non prende in considerazione a priori il processo del doppio crossing over.
Quando si costruisce una mappa è meglio sempre utilizzare più geni possibili separati da distanze brevi.
Marco scusami puoi spiegarmi meglio,nel reincrocio a 3 punti, come hai fatto a stabilire la distanza tra i geni a,b,c?
RispondiEliminaQuesta parte che hai scritto non mi è molto chiara..."Per determinare se sono ricombinanti o parentali queste due classi di geni non importa sapere se i nostri moscerini sono a+ o a.
La distanza a<-->b calcolata come la percentuale di ricombinanti sul numero totale della progenie sarà di 350+338+231+218/5109 x 100= 22,5 u.m
Lo stesso ragionamento che abbiamo applicato per a e b lo applichiamo ad a e c; osservando sempre i dati sopra abbiamo che:
350+338+15+7/5109 x 100= 13,89 u.m
Stesso discorso anche per b e c:
231+218+15+7/5109 x 100= 9,3 u.m".
Grazie anticipatamente,se puoi risp al più presto!!!!!!
Perdonami, credevo di aver corretto il post da orrori di battitura, ho modificato la frase in questione, e aggiungo anche un immagine a breve, inoltre stavo scrivendo proprio ora un post sulla ricombinazione, dov aggiungo qualche notiziola sulla frequenza di ricombinazione...se non ti sarà lo stesso chiaro fammelo sapere...
RispondiEliminaNon riesco a capire come fai a dare: per la distanza a<-->b i seguenti numeri :350+338+231+218;
RispondiEliminapoi per la distanza a<-->c :350+338+15+7;
Infine per b<-->c :231+218+15+7.
Cioè qual è il criterio con cui attribuisci tali valori?
Grazieeee^^
E' più o meno il problema che ti sei posta nell'altro post, quei numeri, 350, 338 ecc...è il numero di individui appartenenti alla progenie del testcross che presentano un determinato genotipo. Come l'altra volta i numeri della progenie li ho inventati di sana pianta solo per fare l'esempio XD.
RispondiEliminaForse mi sono espressa male...Sorry!Il fatto dei num che sono casuali l'ho capito(anche perchè me l'avevi detto appunto nell'altro post!XD)...Non mi è chiaro perchè nel primo caso,a proposito della distanza tra a e b,non hai per esempio considerato il valore 15 e 7(che sono quelli generati da un doppio crossing over),poi xkè nel secondo caso,riguardo la distanza a e c non considerato 231 e 218 e infine nel 3 caso,sulla distanza b e c non hai scritto 350 e 338.
RispondiEliminaera questo il problema che mi stavi ponendo? i numeri da dove li avevo presi? oppure mi stavi chiedendo perchè addizionavo una determinata classe di ricombinanti? fammi sapere! Ora mi fai sentire in colpa, spero di non starti facendo perdere tempo prezioso! Io domani sto piantato davanti al pc, non nessun altr impegno, se hai altri dubbi a riguardo e ti posso aiutare scrivi ti risponderò al più presto!
RispondiEliminaEsatto!!!xkè addizioni solo una determinata classe di ricombinanti?qual è il criterio in base al quale escludi gli altri?
RispondiEliminaScusami tu x il disturbo e nn sentirti sempre in colpa...XD!
Allora, due cose, primo attenta all'immagine in più ch ho messo ho sbagliato la posizione dei geni, credo che si contraddica con quanto dico domani la sostituisco...in risposta al tuo quesito...beh leggi attentamente, ho scritto, ricorreggendo, nel primo commento infatti ti avevo chiesto perdono che per determinare se una classe particolare della progenie è ricombinante o parentale per queste due classi di geni non importa sapere se i nostri moscerini sono c+ o c.
RispondiEliminaquindi osserva l'immagine dove è elencata la progenie del testcross...ci sono vari genotipi, da essi cancella mentalmente c+ e c...nel primo esempio in questo caso 350+338+231+218...prendiamo in considerazione (a+ b)(a b+)e di nuovo (a+ b) e (a b+)...stiamo calcolando la frequenza di ricombinazione (si calcola prendendo in considerazione sempre prendendo in considerazione 2 geni alla volta...) ti è più chiaro?
Mhhh...forse ho capito!Nel primo caso hai considerato solo i genotipi eterozigoti (se così possiamo definirli!)--->a+b e ab+ ;perciò hai escluso i num 17 e 5 che invece hanno ciascuno genotipi omozigoti???
RispondiEliminaMi mostri anche,gentilmente,il secondo e il terzo caso?Hai escluso sempre c e c+?
ps:puoi scrivermi anche domani!
Mi sono confusa...volevo dire,nel secondo caso,x la distanza a e c ,escludo b e b+(e non di nuovo c e c+)???
RispondiEliminaed invece x il terzo caso,ovvero x la distanza b e c, escludo a e a+???
si il ragionamento è quello!!!
RispondiEliminaOk!;) Marco ma il post sulla frequenza di ricombinazione non è ancora pronto o sono io che non riesco a vederlo?
RispondiEliminaCIAOOO
arriva subitissimo
RispondiEliminaScusami Marco...sai cos'è una funzione di mappa?????
RispondiEliminaHo pubblicato un post a riguardo, lo trovi in home page...
Eliminasi,grazie!!!ti ho mandato un'e-mail...
RispondiEliminaChe casino
RispondiEliminadimmi cosa non ti è chiaro XD
RispondiElimina